Example of topic of interest (non restrictive) : BioSonar - new files to upload since 30th sept
One of the possible (non restrictive) topic of interest in this workshop is the biosonar analysis (mostly from bats or dolphins): Which neural processes underlie pulse train emission of biosonar? Are there pulse categories? What is the high / social information content of these sonar, and if any, at which scale? Which could be the efficient decompositions / self-learned representations for these complex sounds?
Some samples and paradigms of sonar sequences of wild dolphin and bats are given below, more are available, please ask to nips4b@gmail.com.
Here a .pdf with the summary of some dolphin data and suggested topic of interest.
First we provide these clear recordings of Stenella dolphin sonar from the Port-Cros National Park, Côte d'Azur [USTV-PELAGOS DECAV project 2011-12]:
The two next files contain sonar of another dolphin species, the biggest one, i.e. Physeter macrocephalus (15 meters, 40T): file c (28MB), and file d (high signal to noise ratio, recorded at Toulon in 2012-DECAV SABIOD, FS=48kHz, 55MB).
Nice 25 minutes of one Physeter have been recorded on 5 channels in Bahamas by NATO, and we have precisely computed the 4D positions of this whale [Glotin 2008] with its real animation on YouTube. The whole recordings 25 minutes x 5 channels at 48kHz and the positions and references are in this archive (500 MB) (here is one sample of 5 min). You find a sparse coding representation of these clicks in [Paris et 2013].

Fig: The sonar sample given below is from Nicky, here with her calf, recorded at Shark Bay Australia (cred. Giraudet 2013).
This Tursiops sonar sample is from the wild dolphin called Nicky, 37 years old, visiting nearly daily Monkey Mia Bay (frequency sample 96kHz, 32 bits, with CR55 hydrophone of Cetacean Research). Here is its time-amplitude representation:

Fig: The time-amplitude representation of this Nicky's sonar short sample (0.7 sec).
Here we give a longer sequence of Nicky (same FS=96kHz, 32 bits, 19MB), recorded at 2m from her noise. You may use Audacity or GNU Octave to read it.
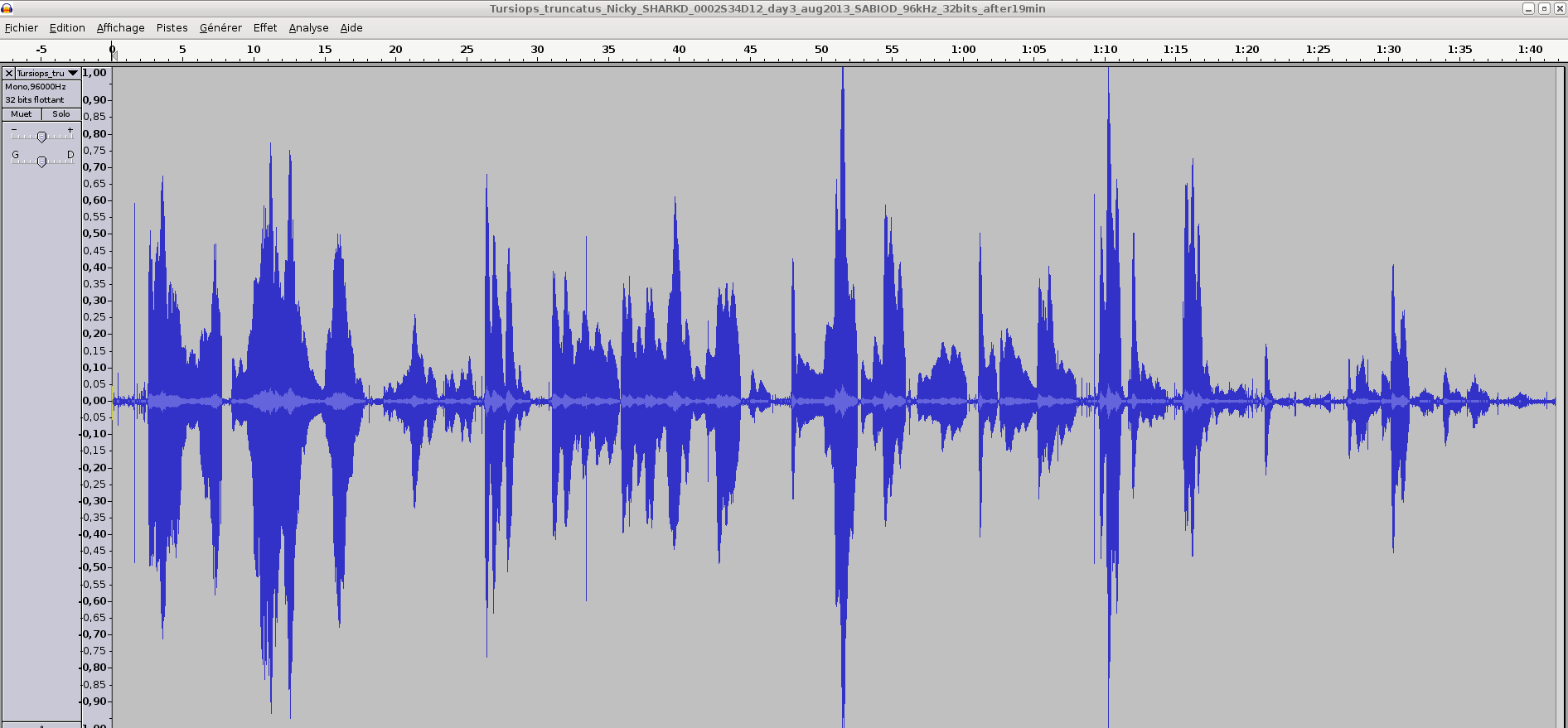
Fig: Longer time-amplitude of Nicky's sonar (100 sec.): same file e.
In [Ryabov 2011] it is shown that Tursiops dolphin are producing the packs of coherent and non-coherent broadband pulses. The waveform and spectrum of coherent pulses are invariable within a pack (see fig. below), but considerably varies from a pack to a pack. The waveform of each non-coherent pulse vary from a pulse to a pulse in each pack, therefore their spectrum also vary from a pulse to a pulse and have many extrema. It is very likely that the non-coherent pulses play a part of phonemes of a dolphin spoken language and the probing signals of dolphin's non-coherent coherent sonar. Efficient feature extraction and classification on sonar sequences are requiered for such studies.

* Sonar of Dolphin River *
We give also Amazone dolphin river (Inia) recorded by David E. Bonnett (bonnettde@gmail.com): Inia, 2007, 96kHz FS, 600Mo , and Inia, 2009, 96kHz FS, 300Mo , and Inia, 2011, 96kHz FS, 300Mo
We also give some Indian River bottlenos dolphins record, recorded by M. Trone (DRS).
Indian River Dolphin, 2013a, 96kHzFS, 358 Mo and Indian River dolphin, 2013b, 500kHz FS, 1Go
From these recordings, you may try to learn features that may correlate with some morphological difference, here are some point of interest on river files.
Similar paradigm applies to bats' sonar: [Kno 2012] shows that if bat echolocation is primarily used for orientation and foraging, it also holds great potential for social communication.
Fig: Bat's sonar share many properties with cetacean one's
- Myopterus bat's sonar sample (credit Cyberio) (frequency sampling=250kHz, duration=4 seconds, 2MB)
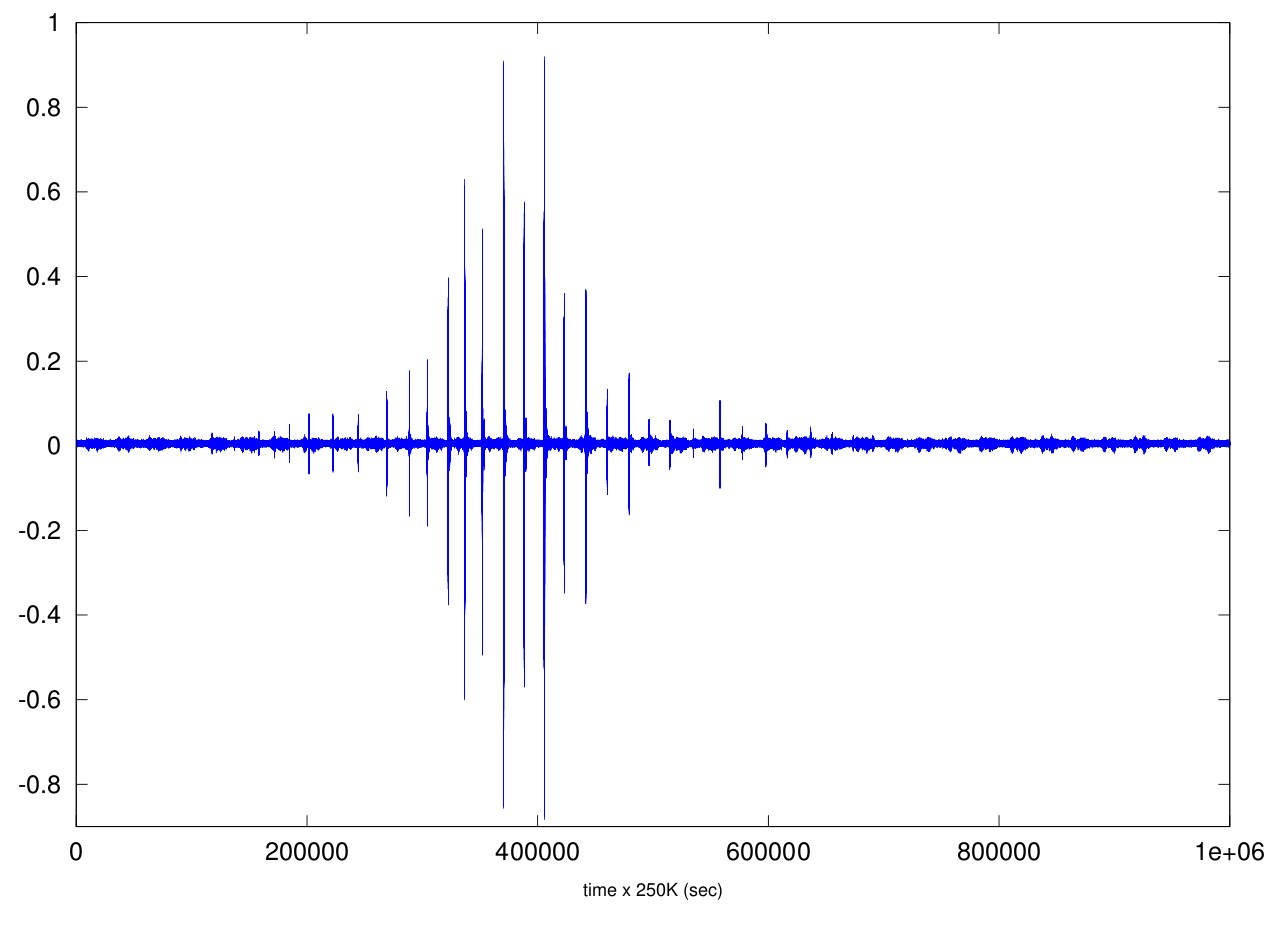
Fig: The time-amplitude representation of this bat sonar sample. The nearest point of approach (NPA) to the microphone corresponds to the highest amplitude (near the sample #400K). Before NPA, the bats flies in direction to the microphone, after NPA the bat emits in the opposite direction.
The communicative function of echolocation calls is still largely unstudied, especially in the wild. The vocal signatures encoding social information in echolocation calls has not been up to now well studied. The authors found pronounced vocal signatures encoding sex and individual identity : free- living males discriminate approaching male and female conspecifics solely based on their echolocation calls. Males always produced aggressive vocalizations when hearing male echolocation calls and courtship vocalizations when hearing female echolocation calls; hence, they responded with complex social vocalizations in the appropriate social context. Advanced statistics may reveal other dependences into biosonar sequences...
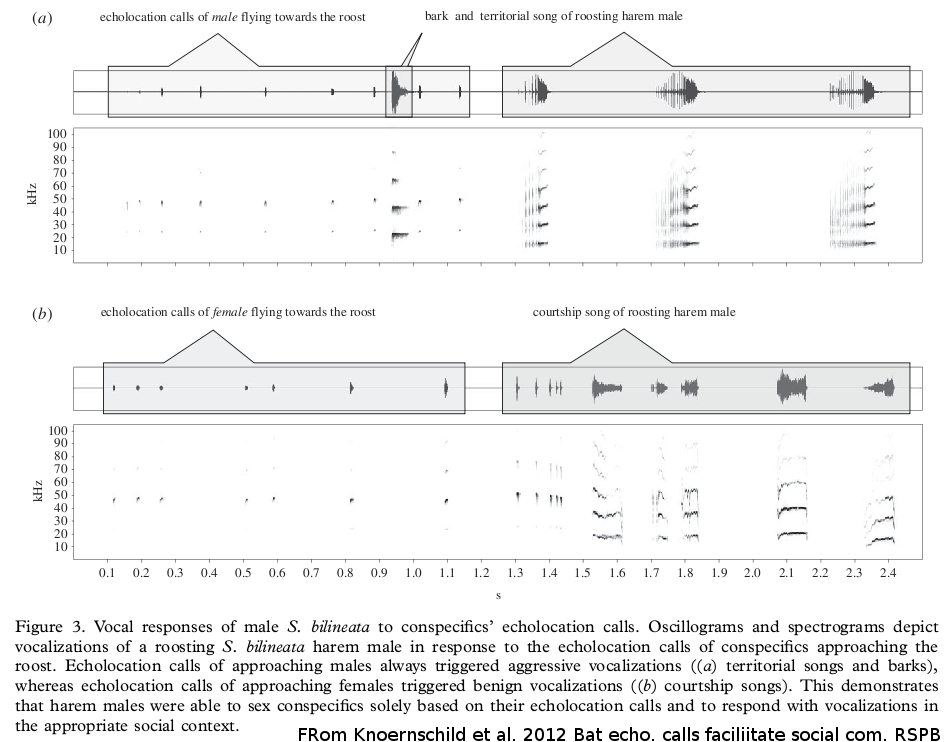
Fig: The sonar sequence dependencies illustration (from [Kno 2012]).
References:
- [Ryabov 2011] Some Aspects of Analysis of Dolphins'Acoustical Signals, Ryabov (2011), Open Journal of Acoustics, 2011, 1, 41-54 doi:10.4236/oja.2011.12006 Published Online (http://www.SciRP.org/journal/oja)
- [Kno 2012] Knörnschild M, Jung K, Nagy M, Metz M, Kalko EKV (2012) Bat echolocation calls facilitate social communication. Proceedings of the Royal Society of London B 279: 4827-4835.
- [Glotin 2008] Glotin H., Caudal F., Giraudet P., 'WHALE COCKTAIL PARTY: REAL - TIME MULTIPLE TRACKING AND SIGNAL ANALYSES', int. Journal Canadian Acoustics, V.36(1), ISSN 0711-6659, DEMO @ sabiod.org
- [Paris 2013] S. Paris and al.(2013) Physeter catodon localization by sparse coding, ICML for Bioacoustics workshop, 2013
- [Benard 2010] Benard F., Giraudet P., Glotin H. (2010) 'Whale 3D monitoring using astrophysic NEMO ONDE 2m wide platform with state optimal filtering by Rao-Blackwell Monte Carlo data association', int. jour. App. Acoustics, (71)

